

2015〕、カナリアカナヘビでは肢あたり最大32個になります〔Baeckens 2015b〕。低木に登る種種は、砂地や岩場などに生息する種類よりも大腿孔の数が少ない傾向があり、これはトカゲのマーキングにおける環境が影響を与えていることを示唆しています〔Baeckens 2015b〕。大腿腺は、ヨロイトカゲ科、クビワトカゲ科、モリイグアナ科、イグアナ科、ツノトカゲ科、ヨルトカゲ科の全属に存在し、アシナシトカゲ科、カメレオン科、フタアシトカゲ科、ドクトカゲ科、トカゲ科、コブトカゲ科、オオトカゲ科の全属に存在しません〔Pianka et al.2003〕。大腿部に存在するために大腿腺という名称がついていますが、ヒョウモントカゲモドキなどのヤモリ科では総排泄孔の頭側に臭腺があり、前肛孔あるいは腹腺とも呼ばれています。オスは腺が発達しており、総排泄腔近位の水平線上に複数並んで位置しています〔Antoniazzi 1993〕。


組織
大腿腺は一列に並んだ独立した小葉/胞状の全分泌腺で、雌雄ともに大腿腺を有しますが、オスの方が顕著に発達しています〔Frye 1991〕。腺は体表皮の胚芽層の陥入によって形成され〔Fergusson 1985〕、真皮層まで伸びており、腺管は拡張して多数の小葉腺/胞腺と融合して腺組織を形成し、腺房は好酸性顆粒で満たされた細胞質を持つ立方体の分泌細胞です〔Frye 1991〕。
内総排泄腔腺
大腿腺は個体間のコミニケーションに使用されていますが、他にも内総排泄腔腺からも化学物質が分泌され、糞便の表面に分泌されます〔Duvall et al.1987,Carpenter et al.1995,López et al.1998,Bull et al.1999,Bull et al.1999,Bull et al.2001,Aragón et al.2000,Wilgers et al.2009,Valdecantos et al.2020〕。異なる供給源から得られる情報化学物質である分泌物は、コミニケーションをとるうえで複数の化学シグナルを生成することができます。例えば、アルジェリアスナカナヘビのメスは、大腿腺分泌物と類似または相補的な情報として糞便中の化学物質を摂取している可能性があり、メスは大腿腺分泌物またはオスの糞便からの化学的手がかりに基づいて、若いオスと縄張り意識を持ったオスを区別することができます〔Nisa-Ramiro et al.2019〕。トッケイヤモリは、皮膚と糞便中の化学物質によって個体識別を行いますが〔Szabo et al.2023〕、オスのホウセキトカゲ(Lioalemus tenuis)の個体識別は糞便中の化学物質に基づいており、臭腺分泌物に基づいているわけではないそうです〔Labra et al.2002〕。
分泌物
オスのカルペタンイワトカゲの大腿腺分泌物は主に脂質とタンパク質で構成されています。これらの脂質の一部はコミュニケーションに重要な役割を果たしている可能性がありますが、タンパク質単独でも少なくとも自己認識が可能です〔Mangiacotti et al.2020,López et al.2005〕。さらにオスは岩の目立つ高い場所に糞をして、複合的な視覚的および化学的シグナルとしても機能します〔López et al.1998〕。大腿腺分泌物と同様に、これらの糞便中に含まれる未解析の化学物質は、性的識別、自己認識、見慣れたオスと見慣れないオスの識別、そしてオスの体格のシグナル伝達を可能にしているそうです〔López et al.1998,Aragón et al.2000〕。オスのサバクイグアナの大腿腺の分泌物は、約80%がタンパク質、20%が脂質成分で構成され、この割合は年間を通して一定です。グリーンイグアナでは、タンパク質構成比も約80%ですが、繁殖期には65%に低下します〔Alberts 1990〕。さらに繁殖期では不飽和脂質をより多く産生され、分泌物の揮発性が高まります〔Albert 1992b〕。グリーンイグアナでにおける大腿腺による個体識別は、性別、血縁レベルまで可能であることが示唆されました〔Alberts 1993b〕。1歳以上のオスのグリーンイグアナでは、大腿腺の生産性、孔の大きさ、そして分泌物中の脂質の割合は、優位個体の血漿テストステロン濃度と正の相関関係にありますが、劣位個体や未成熟のオスでは、未発達です。繁殖期である11月から1月に、分泌物の産生も盛んになります〔Alberts 1992a〕。なお、オスのヒョウモントカゲモドキは実際に舌を動かして分泌物を判別することもします〔Mason 1992〕。
役割
大腿腺の分泌物は、イグアナの周囲の環境に擦り付けられ(マーキング)、他のイグアナに発見されると、種特異的な様々な反応を引き起こします。サバクイグアナと熱帯のグリーンイグアナの比較では、サバクイグアナは紫外線を利用して地上の分泌物を検出し、分泌物の揮発性が低いため、分泌物を舌で直接触れて認識します。一方、熱帯のグリーンイグアナの分泌物は紫外線検出に依存していません。なぜなら、揮発性が高いため分泌物が臭いやすい特徴があるからです。オスはこれらの分泌物を性交渉中の同じオスの戦闘能力を評価するために使用します〔Olsson et al.2003,Martín et al.2006,Martín et al.2015〕 。メスはオスからの求愛はもちろんのこと、質の高い相手を選ぶ要因としても利用します〔Martín et al.2014,López et al.2006〕。

疾病
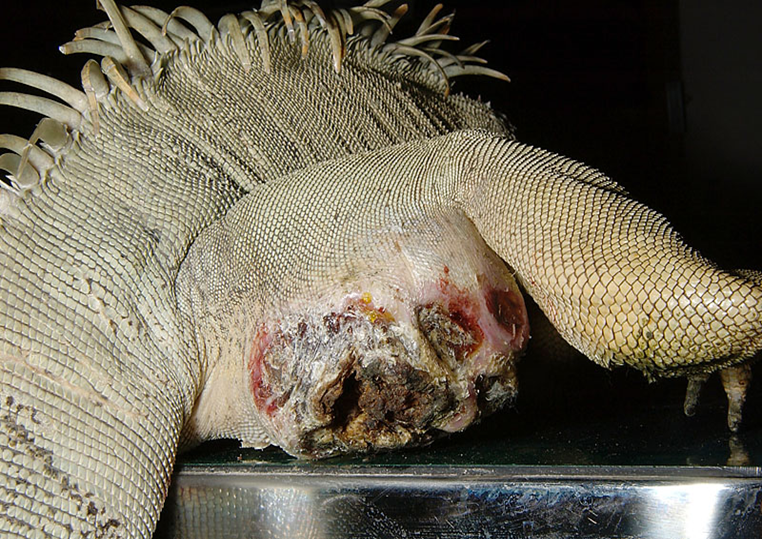
オスのイグアナでは、硬結した分泌物が数個から多数、大腿部に付着している個体を見ることがあります。分泌物は腺の開口部よりもはるかに大きくなっており、周囲に炎症を起こしている場合もあります。これらのイグアナの多くは、塞栓した分泌物を手で押し出す必要があります。炎症などのトラブルを起こすのはイグアナが圧倒的に多く、時にフトアゴヒゲトカゲで発生する程度です。大腿腺の過剰分泌や硬結を予防するには、定期的な温浴が推奨されています。


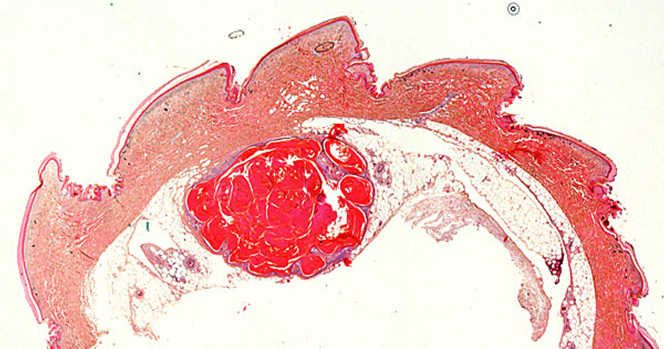
蓄積した分泌物に二次感染が起こることがあります。
炎症が慢性化を起こして、肉芽腫に発展したり、まれに腫瘍の報告があります。

待望の新刊! 爬虫類の病気百科

ポチップ
エキゾチックアニマル臨床の第一人者 霍野晋吉が贈る、獣医師そして飼育者、ブリーダーまで、全爬虫類関係者へ送る医学バイブル
動物看護師の教科書

爬虫類好きなら持っていないといけない
参考文献
- AlbertsAC.Chemical and behavioral studies of femoral gland secretions in iguanid lizards.Brain Behav Evol41(35):255-260.1993a
- Alberts AC,Phillips JA,Werner 01.Sources of intraspecific variability in the protein composition of lizard femoral gland secretions.Copeia3:775-781.1993b
- Alberts AC.Seasonal productivity of lizard femoral glands:relationship to social dominance and androgen levels.Physiol Behav51(4):729-33.1992a
- Albert AC,Sharp TR,Werner 01,Weldon PJ.Seasonal variation of lipids in femoral gland secretions of male green iguanas (Iguana iguana).J Chern Ecol18(5):703-711.1992b
- Alberts AC.Chemical properties of femoral gland secretions in the desert iguana,Dipsosaurus dorsalis.J Chern Ecol16(1):13-25.1990
- Antoniazzi MM,Jared C,Pellegrini CMR,Macha N.Epidermal glands in squamata:morphology and histochemistry of the pre-cloacal glands in Amphisbaena alba (Amphisbaenia).Zoomorphology113:199-203.1993
- Aragón P,López P,Martín J.Size-dependent chemosensory responses to familiar and unfamiliar conspecific faecal pellets by the Iberian rock-lizard,Lacerta monticola Ethology106:1115-1128.2000
- Baeckens S.Chemical signalling in lizards: an interspecific comparison of femoral pore numbers in Lacertidae.Biological Journal of the Linnean Society114:44‐57.2015b
- Baeckens S.Evolution and role of the follicular epidermal gland system in non-ophidian squamates.Amphibia-Reptilia36(3):185‐206.2015
- Bull CM,Griffin CL,Johnston GR.Olfactory discrimination in scat-piling lizards.Behav10:136-140.1999
- Bull CM,Griffin CL.Perkins MV.Some properties of a pheromone allowing individual recognition from the scats of an Australian lizard, Egernia striolata. Acta Ethol2:35-42.1999
- Bull CM,Griffin CL,Bonnett M,Gardner MG,Cooper SJ.Discrimination between related and unrelated individuals in the Australian lizard Egernia striolata.Behav Ecol Sociobiol50:173-179.2001
- Carpenter GC,Duvall D.Fecal scent marking in the western banded gecko (Coleonyx variegatus).Herpetologica51:33‐38.1995
- Duvall D,Graves BD,Carpenter GC.Visual and chemical composite signalling effects of Sceloporus lizards fecal boli.Copeia:1028-1031.1987
- Fergusson B.Hormonal Control of femoral gland secretion in the lizard Amphibolurus omalus.Gen Comp Endocrinol, 57 (3):371-376.1985:508.1991
- Labra A,Escobar CA.Aguilar PM,Niemeyer HM.Sources of pheromones in the lizard Liolaemus tenuis.Rev Chil Hist Nat75:141-147.2002
- López P,Amo L,Martín J.Reliable signaling by chemical cues of male traits and health state in male lizards,Lacerta monticola.J Chem Ecol32:473‐488.2006
- López P,Martín J.Chemical compounds from femoral gland secretions of male Iberian rock lizards,Lacerta monticola cyreni.Z.Naturforschung C60:632‐636.2005
- López P,Aragón P,Martín J.Iberian + lizards (Lacerta monticola cyreni) assess conspecific information using composite signals from faecal pellets.Ethology104:809‐820.1998
- Mason RT.Reptilian pheromones.In Biology of the Reptilia.Gans C,Crews D eds.University of Chicago Press: Chicago IL, USA,Volume18:p114‐228.1992
- Mangiacotti M,Martín J,López P,Reyes-Olivares CV,Rodríguez-Ruiz G,Coladonato AJ,Scali S,Zuffi MAL,Sacchi R.Proteins from femoral gland secretions of male rock lizards Iberolacerta cyreni allow self-But not individual-Recognition of unfamiliar males.Behav.Ecol.Sociobiol74:68.2020
- Martín J,López P.Condition-dependent chemosignals in reproductive behavior of lizards.Horm Behav68:14‐24.205
- Martín J,López P.Pheromones and chemical communication in lizards. In Reproductive Biology and Phylogeny of Lizards and Tuatara; Rheubert JL,Siegel DS,Trauth SE eds.CRC Press: Boca Raton,FL,USA:p43‐77.2014
- Martín J,López P.Vitamin D supplementation increases the attractiveness of males’ scent for female Iberian rock lizards.Proc R Soc Lond B273:2619‐2624.2006
- Mason R.T.Reptilian pheromones.Biology of the Reptilia18:114‐228.1992
- Mayerl C,Baeckens S,Van Damme R.Evolution and role of the follicular epidermal gland system in non-ophidian squamates.Amphib-Reptil36:185-206.2015
- Nisa-Ramiro C,Rodríguez-Ruiz G,López P,da Silva Junior PI.Trefaut-Rodrigues M,Martín J.Chemosensory discrimination of male age by female Psammodromus algirus lizards based on femoral secretions and feces.Ethology125:802-809.2019
- Olsson M,Madsen T,Nordby J,Wapstra E,Ujvari B,Wittsell H.Major histocompatibility complex and mate choice in sand lizards.Proc R Soc Lond. B270 (Suppl. 2), 254–256.2003
- Pianka E,Vitt L.Lizards Windows to the Evolution of Diversity.University of California Press:p92.2003
- Szabo B,Ringler E.Geckos differentiate self from other using both skin and faecal chemicals: Evidence towards self-recognition? Anim.Cogn26:1011-1019.2023
- Weldon PJ.Flachsbarth B,Schulz S.Natural products from the integument of nonavian reptiles.Nat Prod Rep25:738-756.2008
- Wilgers DJ,Horne EA.Discrimination of chemical stimuli in conspecific fecal pellets by visually adept iguanid lizard,Crotaphytus collaris.J.Ethol27:157-163.2009
- Valdecantos S,Ruiz-Monachesi MR,Labra A.Testing the functionality of lipids from feces in the conspecific recognition of the weeping lizard,Liolaemus chiliensis.J Herp54:476- 479.2020