

海洋生活
ウミガメ(上科 Chelonioidea)は、約1億1千万年以上前に他のカメ類から分岐した単一の系統に属する海洋爬虫類であり 、その進化の過程で海洋環境に特化した独自の解剖学的・生理学的適応を発達させてきました〔Lutcavage et al.1997〕 。これらの適応は、広大な海洋での摂餌、長距離移動、深海への潜水、そして陸上での産卵という複雑なライフサイクルを可能にしています 。現生のカメとしては大型の部類であり、最小種のヒメウミガメでも成体になれば甲長が60〜70cmとなり、最大種はオサガメで甲長130〜160cmに達します。
骨格系・外皮系
甲羅と骨格
ウミガメの甲羅は腹側の腹甲と背側の背甲からなる骨甲板で構成されており、内部臓器の保護と支持の役割を果たします 。骨甲板の表面は硬く柔軟な角質甲板で覆われ、甲板の数やパターンは、種の識別に用いられる重要な特徴です 。 しかし、オサガメは他のウミガメとは異なり、骨甲板を持たず、代わりに小さな骨のモザイクで支持された革のような皮膚で覆われています 〔亀崎 1994〕。この柔軟な甲羅は、オサガメが水深1,000m以上、時には1,200mにまで潜水する際の極端な水圧に耐えることを可能にしています 。ほとんどのウミガメ種では脊椎が背甲に融合していますが、オサガメの脊椎は甲羅に融合しておらず、これも深海潜水への適応と考えられます 。 ウミガメの頭部と四肢は、淡水性のカメとは異なり、甲羅の中に完全に引っ込めることができません 。これは流線型の体形と相まって、水中での抵抗を減らし、効率的な遊泳を可能にしています 。
鰭(フリッパー)
ウミガメの四肢の長い指は融合して鰭(フリッパー)を形成しています 。大きく強力な前肢は、水中を8の字を描くように動かすことで推進力を生み出し 、後肢は舵の役割を果たし、方向転換と安定性を提供します 。メスのウミガメは、産卵時に後肢を使って砂浜に卵室を掘るという別の役割も果たします 。

頭部と摂餌適応
ウミガメは歯を持たず、代わりに鋭い嘴状の口を持ち、獲物を砕いたり引き裂いたりするのに適応しています 。種によって口の形状は異なり、食性に合わせて特化しています 。例えば、オサガメの口には後方に向いた棘状の突起(乳頭突起)があり、主食であるクラゲが口から逃げるのを防ぎます 。アオウミガメは海草や海藻を食べるため、繊維質の餌を効率的に摂取できるよう鋸歯状の顎を発達させています 。アカウミガメは、貝類やカニなどの硬い獲物を砕くための大きく強力な顎の筋肉を持っています 。
呼吸器系
ウミガメは爬虫類であるため、水中で呼吸することはできず、空気呼吸を必要とします 。しかし、彼らは水中で長時間息を止める驚くべき能力を持っています。安静時には4〜7時間も潜水することができ、これは心拍数を大幅に遅らせて酸素を節約する能力によるものです 。 ウミガメは体の大きさに比べて肺が比較的大きく、ウミガメ、特にカメ科の種では、主な酸素貯蔵庫は肺となっており、全身に存在する酸素の70%以上を占めています〔Lutz et al.1985,Lutcavage et al.1997〕。これは、哺乳類が血液や組織に酸素を多く貯蔵するのとは対照的です 。さらに、カメ科の種の筋肉のミオグロビン濃度は陸生哺乳類と同程度ですが、潜水哺乳類やペンギンに見られる濃度よりはるかに低いです〔Lutz et al.1985〕。このような特徴により、多くの空気を蓄えることができ、他の爬虫類よりも長く呼吸を止めることができます。 なお、オサガメは、他の小型のウミガメの2倍の血液酸素運搬能力を持ち、血液と組織の酸素貯蔵量が肺とほぼ同等に分かれていると報告されています。ウミガメの肺は、潜水時の主要な酸素貯蔵庫としてだけでなく、また浮力調節器官として二重の機能を持っています 。潜水時には、肺の空気量が圧縮され、浮力が減少します 。
気管の適応
オサガメの気管は、深海潜水に特化した顕著な適応を示します 。幼体のオサガメの気管は、他の爬虫類と同様に円形の軟骨環と結合組織からなる構造ですが 、成体になると、気管はほぼ連続した楕円形の軟骨管に変化します 。この構造は、潜水中に水圧によって気管が容易に虚脱し、深海での圧力関連の問題を回避するのに役立ちます 。 成体の気管の内壁には、広範で密な勃起性血管叢が発達しており、冷たい吸気を温め、加湿する役割を果たします 。これは、哺乳類や鳥類の鼻甲介に相当する機能であり、呼吸による熱損失を防ぐ熱制御に寄与すると考えられています 。
総排泄腔呼吸
ウミガメは主に肺呼吸をしますが、一部の水棲ガメは総排泄腔呼吸(腸呼吸)も行います。総排泄腔呼吸は、総排泄腔から水を取り込み、袋状の器官で酸素を吸収する仕組みです。この呼吸方法は、肺呼吸に比べて酸素の取り込み効率は低いですが、水中での活動を補助する役割があります。特殊な筋肉が甲羅の内外に水を移動させ、この交換を促進します。
代謝率が低い
ウミガメはリラックスしているときや眠っているときは、代謝率を大幅に下げることができます〔Hochscheid et al.2005〕。つまり、酸素の消費量が少なくなり、長時間息を止めておけるのです。これは、エネルギーを節約し、必要なときに水中に留まることができる賢い適応です。
循環器系
ウミガメを含むほとんどの爬虫類は、2つの心房と1つの心室からなる3心室の心臓を持っています 。哺乳類や鳥類とは異なり、爬虫類の肺循環と体循環は完全に分離されていません 。これにより、心臓内で血液が混合する「心臓シャント」が可能となり、生理学的ニーズに応じて肺への血流を調整することができます 。 ウミガメの心拍数は、潜水深度によって大きく変化します。海面にいる時は1分間に約21回ですが、潜水すると急激に減少し、深く潜るほど少なくなり、140メートルを超えると1分間に2回程度になります。
心臓シャントのメカニズムと生理的意義
爬虫類の心室は、筋肉の隆起(muscular ridge)によって部分的に3つの腔(cavum venosum, cavum arteriosum, cavum pulmonale)に分かれています 。これにより、肺から戻る酸素豊富な血液と体から戻る酸素の少ない血液が混合し、肺への血流を迂回させる「右-左シャント(R-Lシャント)」や、肺循環に再循環させる「左-右シャント(L-Rシャント)」が発生します 。
- 右-左シャント(R-Lシャント): 酸素の少ない血液が肺を迂回して体循環に送られるシャントです 。潜水時など、効率的なガス交換の要求が低い状況下で減少すると考えられています 。これにより、肺への血流を制限し、窒素の取り込みを減らすことで、潜水時間を最大化するのに役立つ可能性があります 。
- 左-右シャント(L-Rシャント): 肺から戻る酸素豊富な血液が肺循環に再循環するシャントです 。活動時には肺血流が増加し、末梢組織への酸素供給を促進すると考えられています 。
これらのシャントは、潜水時の心拍数減少(徐脈)や末梢血管収縮といった「潜水反応」と連携して機能します 。ウミガメは、この潜水反応によって心拍数を大幅に低下させ、酸素消費を抑えることができます 。
消化器系
ウミガメの消化管の形態は、その食性と密接に関連しており、種ごとの食料源への適応を反映しています 。
食道
全てのウミガメ種において、食道は管状の筋膜性器官であり、内壁には尖った角質の乳頭突起が胃の方向に向かって並んでいます 。これらの乳頭突起は、食物の摂取を助け、潜水時の圧力変化による逆流を防ぐ機能を持つと考えられています 。オサガメの食道は特に長く、体長のほぼ半分に達します 。
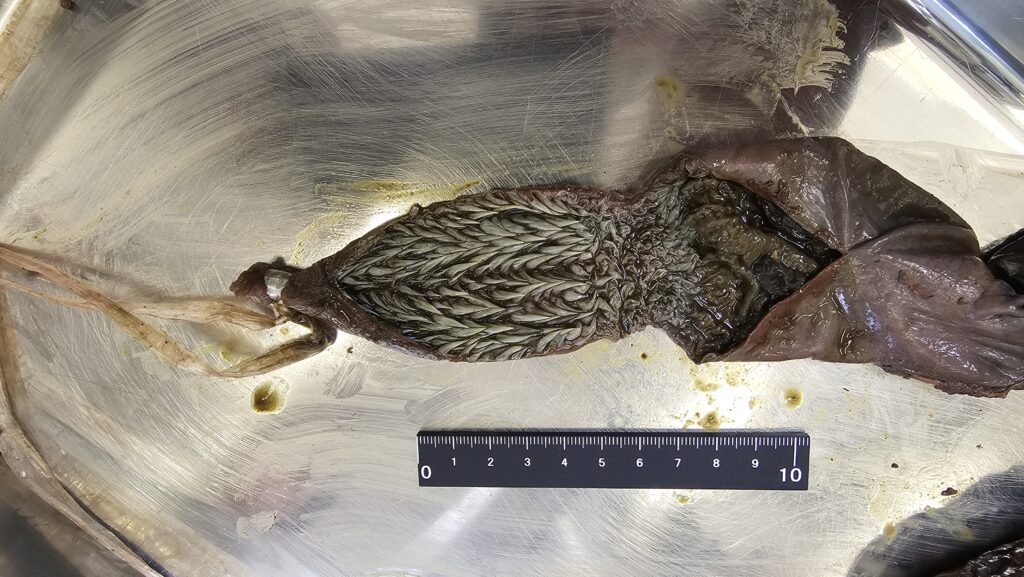
胃
胃は全ての種で体の左側に位置しています 。アオウミガメ、オリーブヒメウミガメ、タイマイの胃はJ字型で、内部には縦方向の粘膜ひだがあります 。アカウミガメの胃は湾曲した管状で、粘膜ひだのパターンに個体差が見られます 。オサガメの胃は曲線的な管状で、粘膜の異なる3つの領域を持ち、それぞれに特化した機能を持つことが示唆されています 。
小腸と大腸
小腸は長く複雑に絡み合った管で、粘膜のひだによって表面積が拡大され、栄養吸収を効率化しています 。大腸は小腸から回盲括約筋を介して移行し、主に水分再吸収の役割を担います 。草食性のアオウミガメでは、大腸の消化における重要性が高く、盲腸(小さな側方嚢)は水分再吸収と植物性炭水化物の細菌による消化に重要です 。
泌尿生殖器系と浸透圧調節
腎臓と尿生成
ウミガメの腎臓は扁平で、腹腔の後壁に密着しています 。爬虫類の腎臓にはヘンレのループが存在しないため、哺乳類のように血漿よりも高浸透圧の尿を生成することはできません 。しかし、アオウミガメは血漿よりもわずかに高浸透圧の尿を生成できると報告されています(最大尿/血漿浸透圧比は約1.2〜1.3) 。尿は尿管から総排出腔またはその派生器官である泌尿生殖洞に排出されます 。
塩類腺
ウミガメは、過剰な塩分を体外に排出するための特殊な「塩類腺」を目の近くに持っています 。これらの腺は、海水を摂取した際に体内に取り込まれる過剰な塩分を、周囲の海水よりも高濃度で排出します 。産卵のために陸に上がったメスガメが「涙を流している」ように見えるのは、この塩類腺からの分泌物であり、感情的なものではなく生理的な機能です 。オサガメは、主食であるクラゲがほとんど海水で構成されているため、他の種よりもはるかに大きな塩類腺を持っています 。
神経系と感覚器官
ウミガメの神経系と感覚器官は、広大な海洋環境での生存とナビゲーションに特化しています。
視覚
ウミガメは水中での視力が優れていますが、水上では近視になります 。彼らの目は、平坦な角膜とほぼ球状のレンズを持ち、主に水中の生活に適応しています 。紫外線や偏光を感知する能力を持つ可能性も示唆されています 。
聴覚
ウミガメは外耳を持たないものの、低周波の音や振動を感知できます 。これは、海岸に打ち寄せる波の音など、沿岸域で一般的に発生する音に敏感であることを意味します 。成体はこれらの音を手がかりに産卵地を特定している可能性も指摘されています 。
嗅覚と化学受容
ウミガメは優れた嗅覚を持っており 、これが産卵のために生まれた浜辺に戻る「帰巣本能」を助けている可能性があります 。また、水中の化学物質(アミノ酸など)を感知し、餌の場所を特定する化学受容能力も持っています 。
磁気受容
ウミガメは、地球の磁場を感知する能力という、人間にはない主要な感覚能力を持っています 。この磁気感覚は非常に洗練されており、方向情報と位置情報の両方を得ることができます 。彼らは磁場の方向をコンパスのように認識し、また地球の磁場の微妙な変化を検出して長距離移動のナビゲーションに利用します 。磁場は、水深、昼夜、天候、季節に関わらず一定であるため、海洋でのナビゲーションに特に有用な情報源となります 。
体温調節(恒温性)
ウミガメは変温動物(ectotherm)であり、体温は主に周囲の環境に影響されます 。彼らは行動や生息地の選択によって体温を調節します 。変温性であるため、代謝率を低下させ、酸素消費量を減らすことで、より長く水中に留まることができます 。
オサガメの特殊な適応
オサガメは、他のウミガメ種とは異なり、体温調節において独自の適応を持っています 。彼らは白色脂肪組織と褐色脂肪組織の両方を含む実質的な脂肪層を持ち 、これにより恒温動物に近い能力を発揮します 。オサガメは水温より数度から最大18℃高い体温を維持でき 、これにより亜極地や深海といったより冷たい水域にも生息範囲を広げることが可能です 。
この体温維持には、以下のメカニズムが寄与しています。
- 巨大体温性(Gigantothermy): オサガメの大きな体サイズは、熱損失を抑えるのに役立ちます 。
- 対向流熱交換器(Counter-Current Heat Exchangers, CCHEs): 後肢に存在するCCHEsは、温かい動脈血と冷たい静脈血の間で熱を交換することで、筋肉で生成された熱を体幹に保持したり、冷たい水中で体幹に熱を伝えたりするのに役立ちます 。
- 行動的体温調節: 熱帯水域ではより冷たい深部に長時間滞在したり、冷たい北部水域では浅い潜水を行ったりすることで、体温を調節します 。
生殖生理
ウミガメの生殖は、陸上での産卵に依存しており、その成功は環境要因に大きく左右されます。
温度依存性性決定(TSD)
ウミガメの性別は、卵の孵化中の周囲の砂の温度によって決定されます 。これは「温度依存性性決定(Temperature-Dependent Sex Determination:TSD)」と呼ばれます 。一般的に、約27.7℃以下の温度で孵化するとオスになり、約31℃以上の温度で孵化するとメスになります 。この閾値間の温度では、オスとメスの両方が生まれます。 地球温暖化による産卵砂浜の温度上昇は、性比をメスに偏らせる可能性があり、将来的な繁殖に不可欠なオス個体の不足を招く懸念があります。
産卵行動と孵化
メスのウミガメは、成熟後2〜6年ごとに産卵のために陸に上がります 。産卵期には、1シーズンに複数回産卵し、1回の産卵で40〜190個(平均約100個)の卵を産みます 。卵は砂の中で45〜75日間孵化し、孵化した子ガメは海に向かって移動します 。
まとめ
ウミガメは、その独特な解剖学的構造と生理学的メカニズムによって、海洋という厳しい環境に適応してきました。柔軟な甲羅と効率的な呼吸器系は深海潜水を可能にし、心臓シャントは潜水中の酸素利用を最適化します。食性に合わせた消化器系の多様な形態、塩類腺による浸透圧調節、そして磁気感覚を含む洗練された感覚器官は、彼らの生存と長距離移動を支えています。
待望の新刊! 爬虫類の病気百科
エキゾチックアニマル臨床の第一人者 霍野晋吉が贈る、獣医師そして飼育者、ブリーダーまで、全爬虫類関係者へ送る医学バイブル

カメを勉強するならコレを聞け
獣医師向けカメセミナーオンデマンド配信
こんなセミナー今までなかった・・・

参考文献
- Hochscheid S,Bentivegna F,Hays GC.First records of dive durations for a hibernating sea turtle.Biol Lett1:82-86.2005
- Lutcavage ME,Lutz PL.Diving physiology.In The Biology of Sea Turtles.Lutz PL,Musick JA eds.Boca Raton.FL.CRC Press:p277-296.1997
- Lutz PL,Bentley TB.Respiratory physiology of diving in the sea turtle.Copeia: 671-679.1985
- 亀崎直樹.オサガメ.日本の希少な野生水生生物に関する基礎資料.水産庁:519-530.1994