

げっ歯類の分類
げっ歯目は、最大かつ最も多様な哺乳類グループであり、全哺乳類種の約40%を占めており、物をかじるのに適した歯と顎を持ち、上顎・下顎の両方に伸び続ける2つの切歯があり、犬歯を欠くことが特徴です。この切歯は物をかじることで次第に削れてゆき、長さを保っており、齧歯目、および学名「Rodentia」はラテン語で「かじる(齧る)」という意味のrodereから来ています〔Crossley 1991,Nowak et al.1983〕。歯は木を削ったり、堅果類の皮をかじったり、身を守ったりするために使用されます。これらの動物は、生息地、体の大きさ、摂食習慣(雑食性から特殊な草食性まで)、食事要件、行動、解剖学的構造、生理学的構造が大きく異なります。げっ歯類は、咬筋の解剖学的および機能的差異に基づいて3つの亜目に分類されます。亜目は、Caviomorpha/Hystrichomorpha(ヤマアラシあるいはテンジクネズミ型/モルモット型齧歯類)〈モルモット、チンチラ、デグーなど〉、Myomorpha(ネズミ型齧歯類)〈マウス、ラット、ハムスター、スナネズミなど〉、およびSciuromorpha(リス型齧歯類)です〈リス、プレーリードッグ、ジリスなど〉〔Capelloet al.2005〕。
歯列と咀嚼の解剖学と生理学
歯列
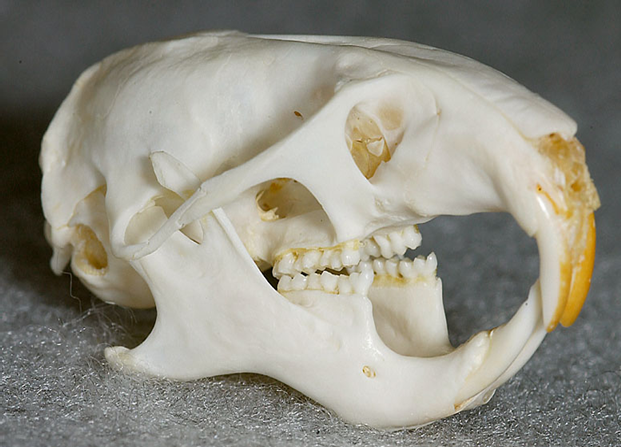
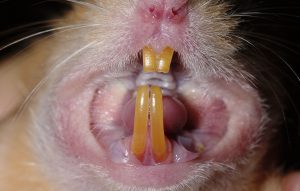
全てのげっ歯類は、この目の最も際立った歯の特徴を備え、発達した上顎および下顎の無根歯の切歯(生涯にわたって継続的に成長および萌出する常生歯で、解剖学的な歯根がありません)を2本を持っています〔Crossley 1991〕。 犬歯を欠き、切歯は歯隙と呼ばれる長い歯のない空間によって小臼歯と分けられています〔Capelloet al.2005〕。ウサギとは異なり、げっ歯類は上顎切歯は単列(1対のみ)です〔Crossley 1991,Capelloet al.2005、Capello et al.2012,Legendre 2003〕。切歯の咬合面はノミ状で、唇側の表面のみがエナメル質で覆われています。ほとんどの齧歯類では、エナメル質は特徴的に黄橙色に着色していますが、これはミネラルが沈着しているためです。モルモット型げっ類では、全ての歯が無歯顎の常生歯になります。また、このグループでは前臼歯と後臼歯が解剖学的に類似しており、一般的に臼歯と呼ばれています〔Crossley 1991,Capelloet al.2005〕。ネズミ型齧歯類とリス型齧歯類の顕著な違いは、ネズミ型げっ歯類の相違は前臼歯がないことで、これは臨床的には意味がありませんが、多様な歯式を生み出します。ネズミ型およびリス型げっ歯類の臼歯は歯根があり、常生歯ではありません。プレーリードッグやその他の草食のジリスの臼歯には複数の歯根を持っています。
| 種類 | 歯式 | 総数 |
| ネズミ、ゴールデンハムスター | 2(I 1/1、C 0/0、P 0/0、M 3/3) | 16 |
| プレーリードッグ | 2(I 1/1、C 0/0、P 2/1、M 3/3) | 22 |
| ハイイロリス | 2(I 1/1、C 0/0、P 1–2/1、M 3/3) | 20~22歳 |
咬筋
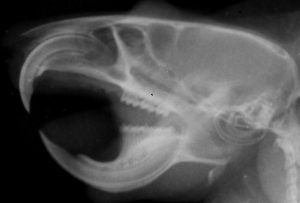
げっ歯類は、他の哺乳類グループと比較して、顎の咀嚼筋群と頭蓋骨の頬骨弓の解剖学的配置が非常に特徴的です〔Cox et al.2011,Cox et al.2012,Druzinsky 2010〕。の3つの咬筋が存在します。咀嚼筋群はいわゆる顎を閉める筋肉で咬筋(浅咬筋、深咬筋、頬骨下顎筋)、側頭筋、内側翼突筋、外側翼突筋からなります〔Cox et al.2012〕。咬筋は頭蓋骨の頬骨弓(人でいうところのほおぼね)と下顎骨の下側面を結ぶ、顎を前に引く筋肉になりますが、げっ歯類の咬筋は他の哺乳類と異なり、頭蓋骨では眼窩(Orbit: 眼球が入るソケット状の窪み)より前に広がり、下顎骨においては前方の臼歯の位置にまで達しており、 これらの特徴は、「かたいものをかじる獣」としての適応と深い関係があります。
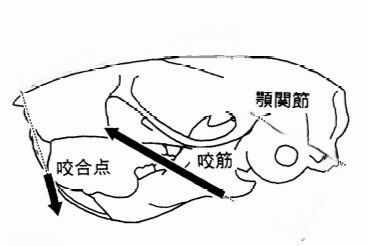
モルモット型げっ歯類は、ネズミ型やリス型と比べて、拡大した咬筋の表層部分が下顎を前後に動かし、縮小した深層部分が顎を内転させて口を閉じるように構成されています〔Crossley 1991,Capelloet al.2005,Cox et al.2012〕。リス型げっ歯類では、深咬筋が吻側まで前方に拡大し、2つの部分(前部と後部)に分かれています。ネズミ型げっ歯類では、咬筋の両部分の起源が吻側まで前方に移動しており、協力して下顎を前後に動かすため、両方の摂食モード(切歯による齧り動作と臼歯による咀嚼動作)に等しく適応しています〔Cox et al.2012,Druzinsky 2010〕。リス型ならびにネズミ型げっ歯類の下顎は吻側尾側への広範囲な運動が可能であり、齧り食い時に吻側に亜脱臼することさえあります〔Crossley 1991,Nowak et al.1983〕。 顎が安静状態のとき、臼歯は咬合しており、下顎は短顎のような外観を呈します〔Crossley 1991,Nowak et al.1983〕。最終的に、切歯が齧り食いし、上下切歯の刃先が接近すると、臼歯の咬合が解除され、食物を咀嚼すると、下顎切歯が上顎切歯の後方に移動し、臼歯が再び咬合します。食物を粉砕する際には、下顎の上方および前方への運動が顕著となります〔Jekl 2009〕。咀嚼筋の解剖は様々ななげっ歯類の異なる食習慣や摂食戦略に反映されます〔Cox et al.2011,Yarto-Jaramillo 2011〕。リ型の深咬筋の吻側部の拡大は、切歯での噛む効率を高めます。逆に、モルモットの深咬筋が縮小しているのは、この種が主に頬歯で噛んで食べ、リスやネズミに比べて切歯で齧る時間が少ないことを示している可能性があります〔Druzinsky 2010〕。ネズミやリスは、主に植物を食べるモルモットよりも硬い食べ物 (ナッツや種子など) を多く消費します。ネズミ目はリス型とモルモット型の解剖学的特徴を融合し、齧りと咀嚼の両方で同等の効率を発揮するため、最も用途が広いです。これら2つの摂食様式は、上顎と下顎の長さの不一致により切歯と臼歯が同時に咬合できないため、互いに排他的です。したがって、これらの動作を行うには前歯列運動が必要です〔Becht 1953,Hiiemae et al.1968〕。
モルモット・チンチラ・デグーの医学 緑書房
他にはない、日本いや世界最高峰の専門医学書・・・

参考文献
- Becht G.Comparative biologic-anatomical researches on mastication in some mammals.Proc K Ned Akad Wet C56:508‐527.1953
- Capello V,Gracis M,Lennox AM.Rabbits and rodents dentistry handbook.Zoological Education Network;Lake Worth.FL;p1‐274.2005
- Cox PG,Jeffery N.Reviewing the morphology of the jaw-closing musculature in squirrels, rats, and guinea pigs with contrast-enhanced microCT.Anat Rec294:915‐928.2011
- Cox PG,Rayfiel EJ,Fagan MJ.Functional evolution of the feeding system in rodents.PLoS One7(4):1‐11.2012
- Crossley DA.Clinical aspects of rodent dental anatomy.J Vet Dent8:131‐134.1991
- Druzinsky RE.Functional anatomy of incisal biting in Aplodontia rufa and sciuromorph rodents—Part 1: masticatory muscles, skull shape and digging. Cells Tissues Organs191:510–522.2010
- Hiiemae K,Ardran GM.A cinefluorographic study of mandibular movement during feeding in the rat (Rattus norvegicus) .J Zool154:139‐154.1968
- Jekl V.Rodents: dentistry.In BSAVA manual of rodents and ferrets.Keeble E,Meredith A eds. British Small Animal Veterinary Association.Gloucester.United Kingdom:p86‐95.2009
- Nowak R,Paradiso J.4th edition. The Johns Hopkins University Press; Baltimore (MD):Walker’s mammals of the world.1983
- .Yarto-Jaramillo E.Respiratory system anatomy,physiology and disease: guinea pigs and chinchillas.Vet Clin Exot Anim14:339–355.2011